
简介
成骨细胞(来自希腊语,ὀστέο-、osteo-和βλαστάνω、blastanō“发芽”)是具有单核的细胞,可合成骨骼。然而,在骨骼形成过程中,成骨细胞以连接的细胞群的形式发挥作用。单个细胞无法制造骨骼。一组有组织的成骨细胞与由一个细胞单位制成的骨骼通常称为骨单位。
成骨细胞是间充质干细胞的特化终末分化产物。它们合成致密、交联的胶原蛋白和数量少得多的特化蛋白质,包括骨钙素和骨桥蛋白,它们构成了骨的有机基质。在不相连的细胞组成的组织群中,成骨细胞产生骨矿物质羟基磷灰石,它以高度受控的方式沉积到有机基质中,形成坚固致密的矿化组织,即矿化基质。矿化骨骼是呼吸空气的脊椎动物身体的主要支撑。它也是生理稳态(包括酸碱平衡和钙或磷酸盐维持)的重要矿物质储存库。

单个成骨细胞
骨骼结构
骨骼是一个大型器官,在呼吸空气的脊椎动物的一生中都会形成和退化。骨骼,通常被称为骨骼系统,作为支撑结构以及维持整个生物体的钙、磷酸盐和酸碱状态都很重要。骨骼的功能部分,即骨基质,完全在细胞外。骨基质由蛋白质和矿物质组成。蛋白质形成有机基质。它被合成,然后添加矿物质。绝大多数有机基质是胶原蛋白,它提供抗拉强度。基质通过羟基磷灰石(别名,羟基磷灰石)的沉积矿化。这种矿物质很硬,并提供抗压强度。因此,胶原蛋白和矿物质一起是一种具有优异抗拉和抗压强度的复合材料,它可以在应变下弯曲并恢复其形状而不会损坏。这称为弹性变形。超过骨骼弹性行为能力的力可能会导致故障,通常是骨折。[需要引证]
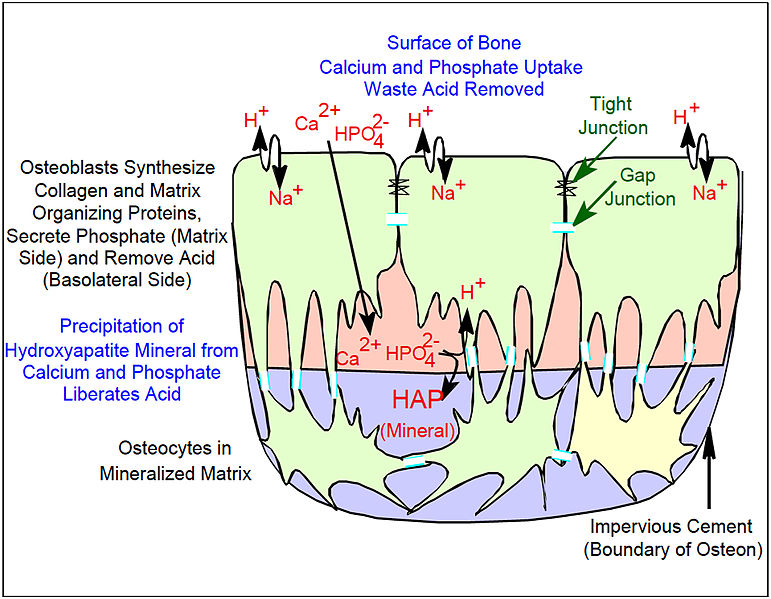
骨形成复合体的主要特征是骨单位,由成骨细胞和骨细胞组成
骨重塑
骨骼是一种动态组织,不断地被成骨细胞和破骨细胞重塑,成骨细胞产生和分泌基质蛋白并将矿物质运送到基质中,而破骨细胞则分解组织。
成骨细胞
成骨细胞是骨骼的主要细胞成分。成骨细胞源自间充质干细胞(MSC)。MSC可产生成骨细胞、脂肪细胞和肌细胞等其他细胞类型。成骨细胞的数量与构成骨髓脂肪组织(MAT)的骨髓脂肪细胞的数量成反比。成骨细胞大量存在于骨膜(骨骼外表面的薄结缔组织层)和骨内膜中。
正常情况下,在呼吸空气的脊椎动物中,几乎所有的骨基质都是由成骨细胞矿化的。在有机基质矿化之前,它被称为类骨质。埋在基质中的成骨细胞被称为骨细胞。在骨骼形成过程中,成骨细胞的表层由立方形细胞组成,称为活性成骨细胞。当骨形成单位没有积极合成骨骼时,表面成骨细胞会变平,称为非活性成骨细胞。骨细胞保持活力并通过细胞突起连接到成骨细胞的表层。骨细胞在骨骼维持中具有重要作用。
破骨细胞
破骨细胞是多核细胞,源自骨髓中的造血祖细胞,造血祖细胞也产生外周血中的单核细胞。破骨细胞分解骨组织,并与成骨细胞和骨细胞一起形成骨骼的结构成分。骨骼内的空腔中还有许多其他类型的骨髓细胞。成骨细胞骨形成所必需的成分包括间充质干细胞(成骨细胞前体)和为骨形成提供氧气和营养的血管。骨骼是一种血管丰富的组织,血管细胞的活跃形成(也来自间充质干细胞)对于支持骨骼的代谢活动至关重要。随着年龄的增长,骨形成和骨吸收之间的平衡趋于负值,尤其是在绝经后女性中,常常导致严重的骨质流失,足以引发骨折,这被称为骨质疏松症。
成骨作用
骨骼由两种过程之一形成:软骨内骨化或膜内骨化。软骨内骨化是从软骨形成骨骼的过程,这是常用方法。这种骨骼发育形式更为复杂:它遵循由软骨细胞形成的第一块软骨骨架,然后被成骨细胞形成的骨骼移除和替换。膜内骨化是间充质的直接骨化,就像在头骨和其他骨骼的膜骨形成过程中发生的那样。在成骨细胞分化过程中,发育中的祖细胞表达调节性转录因子Cbfa1/Runx2。第二个必需的转录因子是Sp7转录因子。骨软骨祖细胞在生长因子的影响下分化,尽管组织培养中分离的间充质干细胞在包括维生素C和碱性磷酸酶底物的允许条件下也可能形成成骨细胞,碱性磷酸酶是一种在矿物质沉积部位提供高浓度磷酸盐的关键酶。
骨形态发生蛋白
软骨内骨骼分化中的关键生长因子包括骨形态发生蛋白(BMP),它在很大程度上决定了软骨细胞分化发生的位置以及骨骼之间的间隙。骨骼替代软骨的系统具有复杂的调节系统。BMP2还调节早期骨骼模式。转化生长因子 β (TGF-β) 是包括 BMP 在内的蛋白质超家族的一部分,BMP 在 TGF β 信号通路中具有共同的信号传导元件。TGF - β 在软骨分化中尤为重要,软骨分化通常在骨形成之前进行,以实现软骨内骨化。另一个重要的调节因子家族是成纤维细胞生长因子(FGF),它决定骨骼元素相对于皮肤的位置。
类固醇和蛋白质激素
许多其他调节系统参与软骨向骨的转变和骨骼的维持。一种特别重要的针对骨的激素调节剂是甲状旁腺激素(PTH)。甲状旁腺激素是在血清钙活性的控制下由甲状旁腺产生的蛋白质。PTH还具有重要的系统功能,包括无论钙摄入量如何,都保持血清钙浓度几乎恒定。增加膳食钙会导致血钙轻微升高。然而,这并不是支持成骨细胞骨形成的重要机制,除非在低膳食钙的情况下;此外,异常高的膳食钙会增加与骨量不直接相关的严重健康后果的风险,包括心脏病发作和中风。间歇性 PTH刺激会增加成骨细胞活性,尽管 PTH具有双功能,并在较高浓度下介导骨基质降解。
骨骼也会为了繁殖以及对营养和其他激素压力的反应而发生改变;骨骼对类固醇(包括雌激素和糖皮质激素)有反应,这些类固醇在繁殖和能量代谢调节中起着重要作用。骨转换涉及合成和降解的大量能量消耗,涉及许多其他信号,包括垂体激素。其中两种是促肾上腺皮质激素(ACTH) 和促卵泡激素。对这些激素和其他几种糖蛋白激素的反应的生理作用尚不完全清楚,尽管 ACTH 可能像 PTH 一样具有双重功能,通过ACTH的周期性峰值支持骨骼形成,但在大浓度下会导致骨骼破坏。在小鼠中,降低肾上腺中 ACTH 诱导的糖皮质激素产生效率的突变会导致骨骼变密(骨硬化骨)。
组织和超微结构
通过电子显微镜在高倍放大下研究保存完好的骨骼,结果表明单个成骨细胞通过紧密连接相连,从而阻止细胞外液通过,形成与一般细胞外液分离的骨隔室。成骨细胞还通过间隙连接相连,间隙连接是连接成骨细胞的小孔,使同一群细胞可以作为一个单位发挥功能。间隙连接还将深层细胞与表层细胞(被骨包围时的骨细胞)连接起来。 通过将低分子量 荧光染料注射到成骨细胞中,并显示染料扩散到骨形成单元中周围和深层细胞,可直接证明这一点。骨骼由许多这样的单元组成,这些单元被称为骨水泥线,它们由没有细胞连接的不透水区隔开。
胶原蛋白和辅助蛋白
骨骼的有机(非矿物质)成分几乎全部是致密的I 型胶原蛋白 ,它形成致密的交联绳,使骨骼具有抗拉强度。成骨细胞通过尚不清楚的机制分泌出定向胶原蛋白层,这些胶原蛋白层与骨长轴平行,每隔几微米就会与骨长轴成直角的胶原蛋白层交替出现。I 型胶原蛋白缺陷会导致最常见的遗传性骨骼疾病,称为成骨不全症。骨的有机基质中分泌有少量但很重要的小蛋白质,包括骨钙素和骨桥蛋白。骨钙素在骨骼以外的地区不会以显著浓度表达,因此骨钙素是骨基质合成的特异性标记物。这些蛋白质将骨基质的有机成分和矿物质成分连接起来。由于这些蛋白质位于矿物质和胶原蛋白之间,因此对于最大基质强度必不可少。然而,在通过靶向破坏相应基因(敲除小鼠)消除骨钙素或骨桥蛋白表达的小鼠中,矿物质的积累并没有受到明显影响,这表明基质的组织与矿物质运输没有显著相关性。
骨和软骨
原始骨骼是软骨,一种无血管的固体组织,其中存在单个软骨基质分泌细胞或软骨细胞。软骨细胞没有细胞间连接,也不以单位为单位协调。软骨由胶原蛋白II型网络组成,该网络由吸水蛋白、亲水性蛋白聚糖保持张力。这是鲨鱼等软骨鱼的成年骨骼。它是更高级动物的 初始骨骼。在呼吸空气的脊椎动物中,软骨被细胞骨所取代。过渡组织是矿化软骨。软骨通过大量表达磷酸盐产生酶而矿化,这会导致钙和磷酸盐局部浓度升高并沉淀。这种矿化软骨不致密也不坚固。在呼吸空气的脊椎动物中,它被用作成骨细胞形成细胞骨的支架,然后被专门降解矿化组织的 破骨细胞去除。
成骨细胞产生一种高级骨基质,由致密、不规则的羟基磷灰石晶体组成,包裹在胶原纤维束周围。这是一种坚固的复合材料,可使骨骼主要形成空心管状。将长骨缩小为管状可减轻重量,同时保持强度。
骨矿化
矿化机制尚不完全清楚。荧光低分子量化合物(如四环素或钙黄绿素)在短时间内给药时会与骨矿物质牢固结合。然后它们在新骨中以窄带形式积聚。这些带贯穿成骨细胞的连续群体。它们发生在狭窄的(亚微米)矿化前沿。大多数骨表面没有新骨形成、没有四环素摄取和没有矿物质形成。这强烈表明,骨形成过程中涉及骨形成群体中协调的促进或主动运输,并且只发生细胞介导的矿物质形成。也就是说,膳食钙不会通过质量作用产生矿物质。骨骼中矿物质的形成机制明显不同于软骨矿化的系统发育上更古老的过程:四环素不会在窄带或特定部位标记矿化软骨,而是会弥漫性地标记,这与被动矿化机制一致。成骨细胞通过紧密连接和受调控的运输将骨骼与细胞外液隔开。与软骨不同,磷酸盐和钙不能通过被动扩散进出,因为紧密的成骨细胞连接隔离了骨形成空间。钙通过协助运输(即通过被动转运蛋白,不会逆梯度泵送钙)在成骨细胞间运输。相反,磷酸盐是由含磷酸盐化合物(包括ATP )的分泌和磷酸酶(磷酸酶裂解磷酸盐以在矿化前沿产生高磷酸盐浓度)共同主动产生的。碱性磷酸酶是一种膜锚定蛋白,是活跃成骨细胞顶端(分泌)面大量表达的特征性标志物。
至少还涉及一个受调控的运输过程。骨矿物质的化学计量基本上是羟基磷灰石在弱碱性pH下从磷酸盐、钙和水中沉淀出来的:
6 羟脯氨酸2−4+ 2 H 2 O + 10 Ca 2+ ⇌ Ca 10 (PO 4 ) 6 (OH) 2 + 8 H +
在封闭系统中,随着矿物质沉淀,酸会积聚,迅速降低pH值并阻止进一步沉淀。软骨没有扩散屏障,因此酸会扩散走,使沉淀继续。在骨单位中,基质通过紧密连接与细胞外液隔开,所以这种情况不会发生。在受控的密封隔间中,只要基质隔间中有钙和磷酸盐,在各种细胞外条件下,去除 H +都会驱动沉淀。酸穿过屏障层的机制仍不确定。成骨细胞有能力通过冗余的 Na/H 交换器 NHE1 和 NHE6 进行Na + /H +交换。 这种 H +交换是酸去除的主要因素,尽管H+从基质空间运输到屏障成骨细胞的机制尚不清楚。
在骨去除过程中,逆向运输机制利用输送到矿化基质的酸来驱动羟基磷灰石进入溶液。
骨细胞反馈
身体活动的反馈可维持骨量,而骨细胞的反馈则限制骨形成单位的大小。另一个重要的机制是骨细胞分泌骨硬化蛋白,骨硬化蛋白隐藏在基质中,是一种抑制维持成骨细胞活性的途径的蛋白质。因此,当骨单位达到极限大小时,就会使骨合成停止。
形态学和组织学染色
苏木精和伊红染色(H&E)显示,由于粗面内质网的存在,活性成骨细胞的细胞质略呈嗜碱性。活性成骨细胞产生大量的I型胶原蛋白。大约10%的骨基质是胶原蛋白,其余为矿物质。成骨细胞的细胞核是球形的,而且很大。活性成骨细胞的形态学特征是明显的高尔基体,在组织学上表现为毗邻细胞核的透明区域。细胞产物大部分被转运到类骨质中,即非矿化基质中。活性成骨细胞可以用I 型胶原蛋白抗体标记,或者使用萘酚磷酸盐和重氮染料固蓝直接显示碱性磷酸酶活性。
热门产品推荐
-

HELA人宫颈癌细胞(STR鉴定正确)
¥1200元
-
HSF人皮肤成纤维细胞(免疫荧光鉴定)
¥1800元
-

Bel-7405人肝癌细胞(STR鉴定正确)
¥1200元
-

Hep G2人肝癌细胞(STR鉴定正确)
¥1200元
-

A549人非小细胞肺癌细胞(STR鉴定正确)
¥1200元
-
H9C2大鼠心肌细胞(种属鉴定正确)
¥1200元
大家都在看
-
hiPSC人诱导多能干细胞培养教程:在培养hiPSC细胞前,需准备好hESC/iPSC完全培养基配制和Matrigel铺板,hiPSC人诱导多能干细胞复苏操作,1. 将水浴锅预热至37℃;并将Matrigel包被的6孔板,提前放置生物安全柜中约1小时恢复···...
阅读详情 -
H9人胚胎干细胞培养条件与方法:1. 将水浴锅预热至37℃,并将Matrigel包被的6孔板,提前放置生物安全柜中约1小时恢复至室温(15~30℃);2. 取4 mL hESC/iPSC完全培养基,按照1:4000比例加入1 μL的hESC/iPSC Supplement C,···...
阅读详情 -
H1人胚胎干细胞培养指南:在培养h1干细胞之前,首先要准备好hESC/iPSC完全培养基配制, 铺板,H1人胚胎干细胞复苏步骤:1.将水浴锅预热至37℃;并将Matrigel包被的6孔板,提前放置生物安全柜中约1小时恢复至室温(15~30℃);···...
阅读详情 -
4T1细胞培养方法:小鼠乳腺癌4T1细胞培养基 90%DMEM+10% FBS+PS,生长条件:气相:95%空气+5%二氧化碳;温度:37℃,4T1细胞形态特征为上皮细胞样,贴壁生长,如果4T1细胞密度达80%-90%,即可进行传代培养,传代方法:1:2至1:3···...
阅读详情 -
常用人肝癌细胞系有哪些及如何选择:目前较为常用的几株人肝癌细胞系SMMC-7721、Bel-7402、MHCC97、HepG2、Hep3B、Huh-7 and PLC/PRF/5,那么人肝癌细胞株如何选择呢?一方面你可以查找相关的文献,另一方面可以考虑选择···...
阅读详情 -
HL-60细胞生长慢解决方法及如何养好:当HL-60细胞传代后生长速度慢且状态不佳时,可竖着培养直到培养基变黄,待细胞密度起来后,状态会有所好转,同时,若HL-60细胞状态很差,可采用半换液的方式:以T25瓶子为例,瓶子里有5m···...
阅读详情







{kind=link}